










































 Ph.D. Equine Nutrition")

Anouk Frieling, MSc Equine Sciences, BSc (Hons)
Horses are seasonal breeders and the breeding season for horses in the northern hemisphere generally lasts from April to October, although for Thoroughbred stallions this can be as early as February. Semen collection for artificial insemination (AI) at most studs starts early on in the year, before the start of the breeding season, and is carried out throughout the breeding season. To perform well as a breeding stallion, whether that be for natural covering or AI, it is important that the diet of the stallion meets their daily requirements to maintain body condition and overall health (Figure 1). The nutrient requirements for a stallion are similar to those of an adult horse, except when actively covering when it is suggested that the nutrient requirements increase about 25%. Like all horses though, requirements can differ between breeding stallions depending on their age, general health, behaviour, breeding frequency during the breeding season and if the stallion is also an active competition horse.

Figure 1. Stallions are seasonal breeders and therefore the breeding season generally lasts from April to October. Due to increased activity the diet should meet the stallion’s requirements during the breeding season.
BODY CONDITION SCORE
It is important for an active breeding stallion to maintain a healthy body weight and body condition throughout the breeding season as it has been shown that an increased body condition in bulls has negative effects on sperm quality and quantity, with sperm motility after collection from heavier bulls being lower, meaning the sperm were less active. Research suggests that the body condition of the pregnant mare has an influence on the pregnancy and the foal. Currently there is limited research available about the effect of body condition of the stallion on fertility, but optimal body condition is known to support the general health of the horse. The body condition of a horse can be scored using body condition scoring (BCS) or fat scoring. It is suggested that a BCS between 5 and 6 is ideal for a stallion during the breeding season, using the 9 point BCS system. The equivalent would be a score of 3 on a scale of 0 to 5. Assessment of fat stores is a helpful tool to manage the energy intake of the horse and will help to avoid over or underfeeding of the stallion.
ENERGY AND PROTEIN REQUIREMENTS FOR STALLIONS
Stallions that do not compete require energy for maintenance during the non–breeding season, whilst during the breeding season the stallion requires energy for maintenance plus additional energy for reproductive activities. Dietary energy is an essential component in the diet to maintain the stallion’s body condition which, as mentioned earlier, is essential for the general health of the stallion. The energy requirements of the breeding stallion during the breeding season depend on the breeding frequency and are similar to the requirements of a horse performing exercise. Inactive breeding stallions (stallions performing a covering every 2 days) have similar energy requirements to those of horses in light work, whilst active breeding stallions (stallions performing two or more coverings per day) require an energy intake comparable to horses performing moderate exercise.
Table 1. The 9-point Body Condition Score system can be used to score the condition of the body of the horse and is a useful tool for managing the diet and overall health of the horse. Stallions during the breeding season should score between 5 and 6 which is between moderate and moderately fleshy and should avoid higher or lower scores which indicate obesity or underfeeding in horses.
Energy is mainly derived from structural carbohydrates (fibre), non–structural carbohydrates (starch and sugar) and fats in the diet. When protein is overfed the excess protein can be used as an energy source by the horse but this is not one of the main energy sources. Fat is a slowly released energy source and therefore provides long-term energy. Non-structural carbohydrates, such as starch, are transformed into quickly accessible energy for immediate use or are stored in the form of glycogen in the horse’s muscle tissue when starch supply exceeds immediate energy demands. The stored glycogen is useful to the horse as it can provide energy when blood glucose levels decrease. About 60 to 70% of the daily energy intake from the stallion’s diet is derived from the fermentation of structural carbohydrates (fibre) into the volatile fatty acids acetate, butyrate and propionate. Therefore, it is important to include high fibre feedstuffs, such as hay, in the stallion’s diet.
Providing a high fibre diet is also essential for a healthy hindgut microbiome composition which aids efficient digestion of nutrients. It is also important that the stallion consumes at least 2.0% of their bodyweight in dry matter. This is equal to at least 10 kilograms of dry matter for a 500kg horse. To meet the energy requirements of the stallion, especially for very active breeding stallions who require more energy for their performance, energy dense cereal grains or oils can be added to the diet. A fibre-based diet supplemented with oils/concentrates supplies the stallion with long-term and short-term energy, helping to maintain ideal body condition and supporting gastrointestinal and mental health throughout the breeding season.
The digestion of dietary protein provides the body with amino acids which are essential for the production of proteins which have different functions in the body of the horse. The mature horse normally does not require high levels of protein in their diet, with recommendations stating that approximately 10% of the diet should consist of protein to meet the maintenance requirements of an adult horse. In contrast, the protein requirements for a stallion during the breeding season changes slightly due to increased activity. Level of activity has an effect on the retention of nitrogen. Stallions that sweat due to increased activity have greater nitrogen losses, leading to a slight increase in dietary protein requirement to compensate for these losses.
Table 2. Energy and protein requirements of a 600 kg breeding stallion depending on breeding frequency (NRC, 2007; Geor et al., 2013)
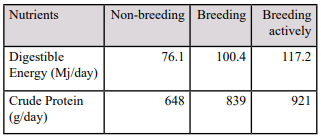
Protein is also important for growth and therefore the young growing stallion will need extra protein in addition to the protein required for maintenance and activity. It is important that daily protein requirements are not exceeded because excess protein will be excreted in the form of ammonia instead of being absorbed by the body which is inefficient use of dietary protein. In table 2 the energy and protein requirements for the adult breeding stallion are shown based on the breeding frequency of the stallion.
VITAMINS AND MINERALS
Vitamins and minerals are only required in small amounts in the diet in comparison to other nutrients, however they have many important functions in the body such as supporting healthy tissue and bone development. Therefore it is important that the diet fed meets the vitamin and mineral requirements of the stallion.
Minerals can be divided into macro minerals and trace minerals. Macro minerals (Calcium [Ca], Phosphorus [P], Magnesium [Mg], Potassium [K], Sodium [Na] and Chloride [Cl]) are required in grams per day whilst trace minerals (Iron [Fe], Copper [Cu], Zinc [Zn], Manganese [Mn], Cobalt [Co], Selenium [Se] and Iodine [I]) are required in milligrams per day. Similarly, vitamins can be divided in fat-soluble (Vitamins A, D, E and K) and water soluble (Vitamins B and C), with fat soluble vitamins able to be stored in the body whilst excess B vitamins and Vitamin C are excreted from the body via the urine.
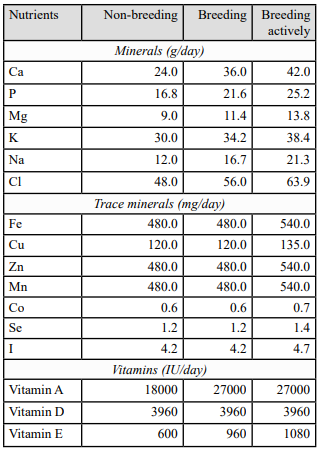
Horses mainly derive minerals and vitamins from the forages and concentrates comprising their daily diet. Vitamin and mineral supplements also offer an option for providing such nutrients and can be particularly useful for stallions who have a tendency to gain weight easily as supplements are generally low energy. Under and overfeeding of minerals and vitamins due to an unbalanced diet can cause various health issues and should therefore be avoided by creating a tailored diet that will suit the nutritional needs of the stallion. For example, a Vitamin E deficiency can result in decreased sperm motility and an increase of abnormal spermatozoa in stallions. Similar to energy and protein requirements, mineral and vitamin requirements of the stallion increase slightly during the breeding season due to their increased activity. Table 3 displays the mineral and vitamin requirements for the adult stallion according to the NRC based on breeding activity.
Table 3. The daily mineral, trace mineral and vitamin requirements for non-breeding, breeding and breeding actively 600 kg breeding stallions according to NRC (2007).
WATER REQUIREMENT FOR STALLIONS
An adult horse requires approximately 4 to 6 litres of water per 100 kg of body weight, although requirements can increase due to environmental temperature and activity. Depending on the intensity and duration of activity, the horse’s water requirements can increase at least 20% as result of water lost in sweat. Requirements can also vary between different housing situations. Stallions that are mainly kept in the stable or are turned out in a sand paddock will require more water in comparison to stallions that have access to fresh forages such as grass which contains approximately 80% water. Providing enough water is essential for digestive purposes but also for body cells that require water to function.
NUTRIENT SUPPLEMENTATION FOR FERTILITY
Semen quality and quantity of natural covering stallions should be as optimal as possible as natural covering stallions can only cover a certain number of mares per breeding season. Optimal quality and quantity results in higher pregnancy rates in mares which increases reproductive performance. Quantity and quality of semen used through AI should also be optimal to be able to cool and freeze the semen for storage and travel.
Techniques have been developed to be able to successfully store semen and inseminate mares. Even though cooling and freezing of the semen improves reproductive efficiency, the process of cryopreservation can also cause oxidative damage. Such damage reduces sperm motility and fertility which has a negative effect on pregnancy rates of inseminated mares. As such, there is an interest in developing methods to decrease semen susceptibility to cold shock.
One of the methods which has been studied to improve stallion fertility is supplementation of different nutrients to optimise metabolic pathways in the body of the stallion. Polyunsaturated fatty acids (PUFAs) are nutrients found in the lipid composition of the semen of the stallion. One of these PUFAs is docosahexaenoic acid (DHA). A study that supplemented the feed of eight fertile stallions with DHA for 14 weeks during a crossover trial. The DHA levels in semen of the supplemented stallions increased by 50%. Results showed that after cooling the sperm for 48 hours the total and progressive motility of the sperm increased. Total motility is the overall percentage of moving sperm, whilst progressive motility is the percentage of sperm that somewhat moves in a straight line. The supplementation of DHA increased the motility of sperm from stallions which had a sperm motility of less than 40% after 24 hours of cooling, prior to supplementation.
Therefore it is suggested that supplementing stallions with DHA has a positive effect on the motion characteristics of semen, which is beneficial for natural covering stallions, and will increase freezability of semen which is beneficial for AI. For more in depth information about PUFAs, DHA and the effect of DHA on reproductive processes, please refer to Nutrient spotlight: essential fatty acids.
Besides DHA, the effects of pomegranate seed oil on stallion fertility has been studied. In a study, 200ml of pomegranate seed oil was supplemented to eight Arabian stallions for a period of 90 days. The results showed no significant effects on fresh collected semen but did show a positive effect on cooled semen, where the pomegranate supplementation increased integrity and viability of semen. Total motility of post-thawed sperm was also increased in stallions supplemented with pomegranate seed oil. The compound L-carnitine has been found in the composition of epididymal secretions of the stallion. Therefore, it is suggested that L-carnitine is able to function as a marker for semen quality in the stallion. Researchers supplemented 20mg of L-carnitine to eight low (progressive motility 50%) stallions for 60 days which resulted in a higher sperm concentration and progressive motility in low fertility horses. Therefore L-carnitine can have a positive effect on sperm motility of stallions producing low motility sperm, making L-carnitine beneficial for sperm production of natural covering stallions and stallions used for AI. The L-carnitine supplementation did not affect semen of the high motility stallions.

Figure 2. Supplementing the stallions feed with nutrients such as DHA and vitamin E has been shown to have positive effects on their fertility.
Research has also focussed on the effects of antioxidant supplementation on stallion fertility. One of the antioxidants that has been studied is Vitamin E. Researchers supplemented 3000IU of vitamin E per day to 15 stallions for 14 weeks and found that supplementation of the antioxidant improved the total and progressive motility of sperm which had been cooled for 48 hours and had been warmed up afterwards for 10-30 minutes.
Further research evaluated the effect of supplementing a combination of DHA derived from micro-algae, Vitamin E and Selenium to stallions. Results of this study suggest that the combination of these nutrients improves the motility of fresh and also cooled sperm after 60 day supplementation, with this benefit being considerable for sperm collected from stallions with a progressive motility of less than 40% which were classified as ‘poor coolers’ as the supplement considerably increased total and progressive motility of stallion sperm.
The results of studies mentioned above show that the supplementation of certain nutrients can enhance fertility of stallions through improved sperm motility and ability to cool and freeze sperm, and by limiting oxidative damage after using these storage techniques. Such benefits may also apply to natural covering stallions as the greater sperm motility will increase their fertility which has the potential to translate to increased pregnancy rates. Limiting oxidative stress after sperm storage for AI increases the insemination success rate in mares with the potential to result in higher pregnancy rates.
SUMMARY
The stallion is a seasonal breeder and is mainly active during the breeding season which for most stallions lasts from April until October. During the breeding season the nutrient requirements of the stallion will increase depending on the age, general health, behaviour, breeding frequency during the breeding season and if the stallion is also an active competition horse. It is important that the stallion maintains a healthy body condition score (score 5 to 6 on a 9-point scale). To achieve this body condition the stallion must be provided with enough energy in their diet, with this energy being derived from structural and non-structural carbohydrates and fats. In addition to energy, protein, minerals, and vitamins, should be balanced as they have important functions within the body, especially for the production of healthy sperm. Supplementing specific nutrients can have positive effects on the fertility of the stallion which can lead to higher pregnancy rates and benefit the breeder. Vitamin E and DHA in particular have been shown to positively impact the semen of the stallion.
