










































 Ph.D. Equine Nutrition")

Anouk Frieling, MSc Equine Sciences, BSc (Hons)
Since domestication, the horse has developed into an incredible athlete being able to perform in diverse disciplines ranging from show jumping to endurance riding (Waller & Lindinger, 2010). Due to this development, the energy requirements of the horse have increased to enable it to perform optimally in each discipline (Harris, 1997). Energy is provided in the diet from fats, carbohydrates and protein, although the latter would only be a secondary source. Dietary energy sources provide the body with fuel to maintain normal biological functions such as growth and repair. In performance horses there is also a large amount of energy needed for muscle contraction (Treiber et al., 2008), making energy a key consideration for performance horse diets. As the outdoor competition season has started, this article will explain the different types of energy sources that can be provided through the diet and how these different sources support optimal performance for different types of exercise.
DIETARY ENERGY SOURCES
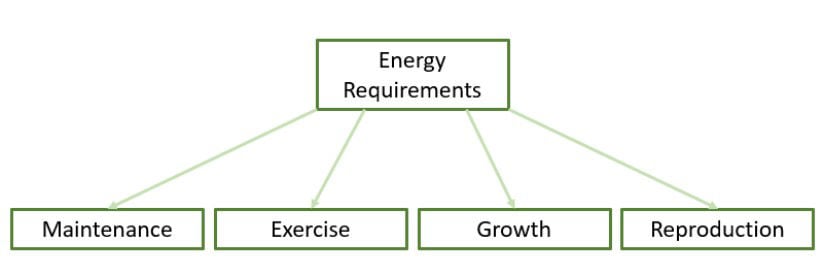
Energy is required for body maintenance, work, growth and reproduction (breeding, lactation) (Figure 1). For the performance horse it is important to have good body condition as they should not be carrying excess weight but should have energy stored in the body that can be used during exercise (Nielsen, 2013). Therefore, to estimate energy requirements, the exercise level of an equine athlete has to be taken into account when composing a diet.
Figure 1. Dietary energy is required for body maintenance, performing exercise, growth of the young horse and for reproduction. The daily energy requirement of the individual horse depends on the intensity of these four factors.
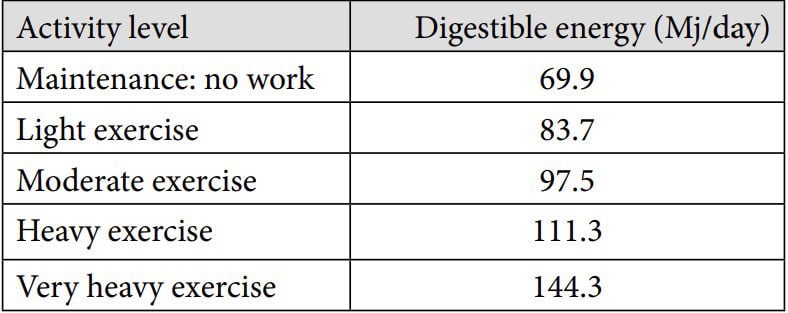
A horse performing high intensity exercise requires more energy in comparison to horses performing lower intensity exercise (Table 1). Dietary energy can be derived from various feedstuffs such as roughage and concentrates and in the UK is expressed in megajoules (Mj) of digestible energy (DE).
Table 1. Energy (Mj/DE per day) requirements depending on exercise level based on a 500kg horse (NRC, 2007)
Dietary fats are commonly supplemented to the diet due to their ability to support and improve coat quality and shine. Besides their supportive function, fats are also energy dense nutrients which can be fed alongside carbohydrates to meet the increased energy requirement of the athletic horse (Warren & Vineyard, 2013). Fats are long-term energy sources, even though they are digested and absorbed fairly quick in the small intestine, fat metabolismin the body is rather slow and therefore energy is slowly released from fats. Because fats are long-term energy sources they can deliver energy during exercise (Potter & Gibbs, 2011) specifically during low-to moderate intensity exercise (Warren & Vineyard, 2013). Dietary fats can be included by feeding plant-based oils such as linseed oil or algae derived oils.
Carbohydrates can be divided into structural and non-structural carbohydrates. Structural-carbohydrates, also known as fibre, are utilised in the horses body through microbial fermentation in the hindgut into the volatile fatty acids (VFA’s) acetate, propionate and butyrate (Richardson & Murray, 2016). Fermentation of fibre, from roughage like hay, in the hindgut provides enough energy to cover the energy requirements for horses performing no or low-intensity exercise (Harris et al., 2017), however supplementation of fats and non-structural carbohydrates, in the form of concentrates, is often required for horses performing moderate or high-intensity exercise to meet their energy requirements (Jose-Cunilleras et al., 2002). The feeding of high-starch diets often receives bad press as previous research analysing a high-fibre and high-starch diet concluded that fibre supports the microbial composition in the hindgut, digestive health and efficiency, whereas high-starch diets can cause ulcers (Andrews et al., 2017), and also alter the microbial population and composition which can result in colic or behavioural changes (Durham, 2009;Destrez et al., 2015). High-starch feedstuffs however do provide sources of quick release energy, giving them a role in the diet if fed at appropriate levels.
Non-structural carbohydrates are mainly derived from concentrates and fresh grass in the form of starch and sugars (Longland & Byrd, 2006). In the body starch and sugars are transferred into glucose, a quickly accessible form of energy ready for immediate use by the body. Excess glucose is stored in muscle tissue in the form of glycogen which is able to provide energy in case blood glucose levels decrease (Potter & Gibbs, 2011).
Protein is a key nutrient for muscle development and growth of the horse. Approximately 10-15% of the total body mass of the horse consists of protein with the muscles containing the largest part of the total percentage (Urschel & Lawrence, 2013). Dietary protein, which is mainly derived from forages and concentrates, consist of amino acids that are necessary for protein synthesis in the body (Urschel & Lawrence, 2013). Protein can be used as an energy source by the body but this only happens if the diet does not contain enough carbohydrates and fats to fulfil energy requirements (Johnson & Duberstein, 2010). Previously it was assumed that excess protein was used as an energy source but more recent research explained that excess protein is excreted in the form of urea in urine. Therefore, this article will focus on carbohydrates and fats as energy sources (Figure 2).
Figure 2. Nutrient pools after absorption used for energy production in the body.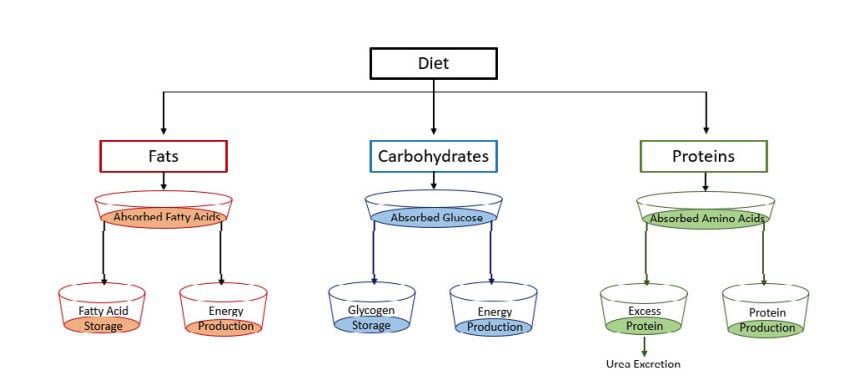
ENERGY METABOLISM FED AND FASTED STATE
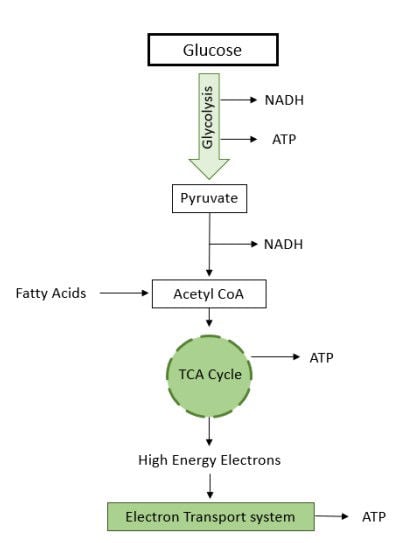
Once carbohydrates and fats are digested and absorbed by the body the mitochondria, organelles in body cells, start to produce energy-carrying molecules (Bertram et al., 2006), of which adenosine triphosphate (ATP) is the main energy molecule in the body (Jonckheere et al., 2012). Energy in the form of ATP is either used immediately, which is called the fed state, or converted for storage by the body (Aird et al., 2018). When nutrients are no longer available in the blood stream during intense exercise the body enters the fasted state and requires energy from the stored nutrients in body cells (Aird et al., 2018). The primary source for energy synthesis in cells is glucose (Figure 3) (Bonora et al., 2012), mainly derived from dietary carbohydrates (Nafikov & Beitz, 2007). Besides glucose, oxidation of fatty acids also produces ATP (Figure 3) (Turner et al., 2014). Oxidation, the process that requires oxygen to produce energy (Longo et al., 2016), of fatty acids creates the high energy molecules NADH + H+ and FADH2 which are used for ATP production (Turner et al., 2014). Full oxidation of fatty acids produces larger amounts of ATP molecules in comparison to that of glucose, although this process requires greater amounts of oxygen. As such, fatty acids produce more energy than carbohydrates but require more oxygen per mole of ATP produced (Turner et al., 2014).
Figure 3. The three processes involved with ATP synthesis. The first process is transferring glucose from the diet into pyruvate which reacts with coenzyme A before entering the TCA cycle. The TCA cycle produces high energy electrons which are involved with the high quantity of ATP (Silverthorn, 2015).
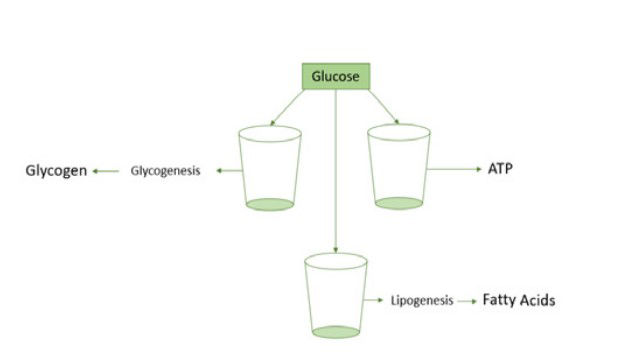
Nutrients that are not immediately used for ATP production are stored in body cells and are used once the horse performs heavy exercise (Harris & Schott, 2013). Excess fatty acids are converted into triglycerides through the process of lipogenesis which mainly occurs in adipose tissue but can also occur in the liver, heart, muscles and pancreas (Saponaro et al., 2015). Excess glucose is converted into glycogen, a glucose polymer (Adeva-Andany et al., 2016), and stored in liver and muscle cells through the process of glycogenesis which is generated by the release of the hormone insulin by the pancreas (Rui, 2014). If there is still excess glucose after energy synthesis and storage in the form of glycogen, glucose is transferred into fatty acids through de novo lipogenesis (Schutz, 2004), and can therefore be stored in adipose tissue (Figure 4) (Ameer et al., 2014).
Figure 4. Once carbohydrates are digested into glucose they can either be used for energy production or stored in body cells. If glucose could be divided into cups, one cup would be used for energy production. When this cup is full another cup would be used for storage of glycogen. If after glycogen storage there is still excess glucose in the blood stream, then glucose would be converted into fatty acids which are stored in adipose tissue.
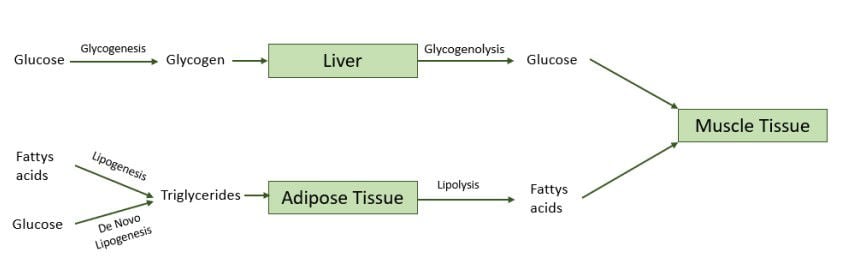
Glycogen and triglycerides are converted into glucose and fatty acids once the horse proceeds into the fasted state during exercise and requires energy from stored nutrients as they cannot eat to produce energy (Figure 5) (Polaket al., 2008; Han et al., 2016). Glycogen is converted through glycogenolysis, a process that does not necessarily require oxygen and therefore glucose can be released quickly during intense muscular activity when oxygen is scarce (Müller et al., 2012). Although the anaerobic metabolism (metabolism without oxygen) of glucose can be useful during exercise, the process also creates lactic acid and yields less ATP compared to aerobic metabolism (metabolism with oxygen) of fatty acids and glucose (Harris & Schott, 2013). Trough the lipolysis process triglycerides are converted into fatty acids (Richard et al., 2000). Fatty acids are always metabolised through aerobic mechanisms, therefore lipolysis requires oxygen which results in a slower release of fatty acids in the blood stream, in comparison to glucose, during intense exercise, making it a long-term energy source (Harris & Schott, 2013).
Figure 5. Metabolic processes utilising dietary energy sources and energy stored within the body to provide muscles with energy during exercise.
FEEDING THE EQUINE ATHLETE
As previously discussed carbohydrates and fats are metabolised through different pathways which creates either short-term or long-term energy supplies for the horse. Due to the difference in either short or long-term energy supply the discipline in which the equine athlete performs should be taken into account when formulating a diet. Endurance horses for example require slow releasing energy sources, which can be provided through dietary fats and a high-fibre diet, as endurance horses exercise for prolonged periods (Harris & Schott, 2013). Roughage, a fibrous feedstuff, is also able to retain fluids and electrolytes which are especially important during prolonged exercise such as endurance (Harris, 2009). Non-structural carbohydrates in the form of concentrates can be supplemented to maintain body weight and to make sure other nutrients requirements are met (Harris & Schott, 2013). Opposed to endurance horses, race horses perform intense exercise within shorter periods and require a combination of quick and slow releasing energy to deliver energy for muscle contraction during this form of exercise (Nielsen, 2013). Therefore, a diet based on roughage supplemented with non-structural carbohydrates, will be able to supply this type of energy release. To meet the energy requirements and maintain body condition, without feeding excessive amounts of starch, fats can be supplemented to the diet (Nielsen, 2013).
Overfeeding carbohydrates or fats can results in constantly triggering the pancreas to release insulin (Wilcox, 2005; Warren & Vineyard, 2013). This constant release can result in insulin insensitivity or resistance, meaning that the cells do not respond properly to the release of insulin and are unable to store all the excess glucose circulating in the blood stream (Kaczmarek et al., 2016). Insulin resistance is associated with health issues such as Equine Metabolic Syndrome (EMS) and laminitis. For more information about insulin resistance and EMS, please refer to Equine metabolic syndrome, or for further information on laminitis see Update on laminitis.
Due to the quick or slow energy release of fats and carbohydrates, the feeding time before exercise should also be taken into account as energy needs to be available when the muscles require it during exercise. Previously Pagan & Harris (1999) explained that feeding grain should be avoided prior to exercise as it was shown to affect glycaemic response and reduced the availability of fatty acids. Because of this response, fatigue occurs earlier when horses are exercised shortly after consuming concentrates (Harris & Harris, 2005). Brunner et al. (2015) found that feeding roughage 2-4 hours prior to exercise to show jumping horses had a positive significant effect on the blood parameters glucose, lactate, insulin, triglycerides and free fatty acids, whilst concentrate feeding did not have a significant effect. Therefore, research has shown that feeding time and energy source are important to consider so that the horse is able to perform optimally.
SUMMARY
For the performance horse the energy sources in the diet are carbohydrates and fats. These nutrients can be included in the diet by providing the horse fibrous feeds like hay, concentrates or plant-based oils like linseed oil. Once these nutrients are digested and absorbed by the body, they will undergo metabolic processes that will convert them into ATP which is an energy carrying molecule. Energy can be immediately used by the body for example for maintenance or light exercise but excess energy can also be stored in liver cells, muscle cells or adipose tissue. By storing excess energy, the body can provide its own energy if the horse is exercising intensively and is not able to consume feed. Energy sources are either released quickly or slow and therefore the discipline of the horse should be taken into account when composing a diet as performing in different disciplines require different types of energy. Besides the type of energy source, the feeding time also has an effect on the energy availability during exercise and should therefore also be taken into account. Therefore, for the athletic performance horse it is important that the diet contains enough energy to maintain body condition and is adjusted to the workload and discipline to provide energy that is required during exercise.
REFERENCES
Adeva-Andany, M. M., González-Lucán, M., Donapetry-García, C., Fernández-Fernández, C., & Ameneiros-Rodríguez, E. (2016). Glycogen metabolism in humans. BBA Clinical, 5: 85-100.
Aird, T. P., Davies, R. W., & Carson, B. P. (2018). Effects of fasted vs fed-state exercise on performance and post-exercise metabolism: A systematic review and meta-analysis. Scandinavian Journal of Medicine and Science in Sports, 28(5): 1476-1493.
Ameer, F., Scandiuzzi, L., Hasnain, S., Kalbacher, H., & Zaidi, N. (2014). De novo lipogenesis in health and disease. Metabolism: Clinical and Experimental, 63(7): 895-902.
Andrews, F. M., Larson, C., & Harris, P. (2017). Nutritional management of gastric ulceration. Equine Veterinary Education, 29(1): 45-55.
Bertram, R., Gram Pedersen, M., Luciani, D. S., & Sherman, A. (2006). A simplified model for mitochondrial ATP production. Journal of Theoretical Biology, 243(4): 575-586.
Bonora, M., Patergnani, S., Rimessi, A., de Marchi, E., Suski, J. M., Bononi, A., Giorgi, C., Marchi, S., Missiroli, S., Poletti, F., Wieckowski, M. R., & Pinton, P. (2012). ATP synthesis and storage. Purinergic Signalling, 8(3): 343-357.
Brunner, J., Liesegang, A., Weiss, S., & Wichert, B. (2015). Feeding practice and influence on selected blood parameters in show jumping horses competing in Switzerland. Journal of Animal Physiology and Animal Nutrition, 99(4): 684-691.
Destrez, A., Grimm, P., Cézilly, F., & Julliand, V. (2015). Changes of the hindgut microbiota due to high-starch diet can be associated with behavioral stress response in horses. Physiology and Behavior, 149: 159-164.
Durham, A. E. (2009). The Role of Nutrition in Colic. Veterinary Clinics of North America - Equine Practice, 25(1): 67-78.
Han, H. S., Kang, G., Kim, J. S., Choi, B. H., & Koo, S. H. (2016). Regulation of glucose metabolism from a liver-centric perspective. Experimental and Molecular Medicine, 48(3): 1-10.
Harris, P. (1997). Energy sources and requirements of the exercising horse. Annual Review of Nutrition, 17: 185-210.
Harris, P. A., Ellis, A. D., Fradinho, M. J., Jansson, A., Julliand, V., Luthersson, N., Santos, A. S., & Vervuert, I. (2017). Review: Feeding conserved forage to horses: recent advances and recommendations. Animal, 11(6): 958-967.
Harris, P. A., & Harris, R. C. (2005). Ergogenic potential of nutritional strategies and substances in the horse. Livestock Production Science, 92(2): 147-165.
Harris, P. (2009). Feeding Management of Elite Endurance Horses. Veterinary Clinics of North America - Equine Practice, 25(1): 137-153.
Harris, P. A., & Schott, H. C. (2013). Chapter 14 - Nutritional management of elite endurance horses. In: Geor, R.J., Harris, P.A., & Coenen, M., (Eds.). Equine Applied and Clinical Nutrition. Saunders Elsevier: China.
Johnson, E. L., & Duberstein, K. J. (2010). How to Feed a Horse : Understanding Basic Principles of Horse Nutrition. University of Florida, IFAS Extension, 2010(2): 1-5.
Jonckheere, A. I., Smeitink, J. A. M., & Rodenburg, R. J. T. (2012). Mitochondrial ATP synthase: Architecture, function and pathology. Journal of Inherited Metabolic Disease, 35(2): 211-225.
Jose-Cunilleras, E., Hinchcliff, K. W., Sams, R. A., Devorand, S. T., & Linderman, J. K. (2002). Glycemic index of a meal fed before exercise alters substrate use and glucose flux in exercising horses. Journal of Applied Physiology, 92(1): 117-128.
Kaczmarek, K., Janicki, B., & Głowska, M. (2016). Insulin resistance in the horse: A review. Journal of Applied Animal Research, 44(1): 424-430.
Longland, A. C., & Byrd, B. M. (2006). Pasture nonstructural carbohydrates and equine laminitis. Journal of Nutrition, 136(7): 2099-2102.
Longo, N., Frigeni, M., & Pasquali, M. (2016). Carnitine transport and fatty acid oxidation. Biochimica et Biophysica Acta - Molecular Cell Research, 1863(10): 2422-2435.
Müller, M., Mentel, M., van Hellemond, J. J., Henze, K., Woehle, C., Gould, S. B., Yu, R.-Y., van der Giezen, M., Tielens, A. G. M., & Martin, W. F. (2012). Biochemistry and Evolution of Anaerobic Energy Metabolism in Eukaryotes. Microbiology and Molecular Biology Reviews, 76(2): 444-495.
Nafikov, R. A., & Beitz, D. C. (2007). Carbohydrate and lipid metabolism in farm animals. Journal of Nutrition, 137(3): 702-705.
Nielsen, B. D. (2013). Chapter 13 - Practical considerations for feeding racehorses. In: Geor, R.J., Harris, P.A., & Coenen, M., (Eds.). Equine Applied and Clinical Nutrition. Saunders Elsevier: China.
NRC. (2007). Nutrient Requirements of Horses, Nutrient Requirements of Horses. Pagan, J. D., & Harris, P. A. (1999). The effects of timing and amount of forage and grain on exercise response in thoroughbred horses. Equine Veterinary Journal, 30: 451-457.
Polak, J., Bajzova, M., & Stich, V. (2008). Effect of exercise on lipolysis in adipose tissue. Future Lipidology, 3(5): 557-572.
Potter, G. D. & Gibbs, P. G. (2011) Feeding the Performance Horse. Texas a & M University Department of Animal Science Equine Sciences Program.
Richard, A. J., White, U., Elks, C. M., & Stephens, J. M. (2000). Adipose Tissue: Physiology to Metabolic Dysfunction. Endotext.
Richardson, K., & Murray, J. A. M. D. (2016). Fiber for Performance Horses: A Review. Journal of Equine Veterinary Science, 46: 31-39.
Rui, L. (2014). Energy metabolism in the liver. Comprehensive Physiology, 4(1): 177-197.
Saponaro, C., Gaggini, M., Carli, F., & Gastaldelli, A. (2015). The subtle balance between lipolysis and lipogenesis: A critical point in metabolic homeostasis. Nutrients, 7(11): 9453-9474.
Schutz, Y. (2004). Dietary fat, lipogenesis and energy balance. Physiology and Behavior, 83(4): 557-564.
Shimazu, T. (1981). Central nervous system regulation of liver and adipose tissue metabolism. Diabetologia, 20(1): 343-356.
Silverthorn, D. U. (2015). Human Physiology: An Integrated Approach, Global 7th Edition. Pearson, Londen, UK.
Treiber, K. H., Geor, R. J., Boston, R. C., Hess, T. M., Harris, P. A., & Kronfeld, D. S. (2008). Dietary energy source affects glucose kinetics in trained arabian geldings at rest and during endurance exercise. Journal of Nutrition, 138(5): 964-970.
Turner, N., Cooney, G. J., Kraegen, E. W., & Bruce, C. R. (2014). Fatty acid metabolism, energy expenditure and insulin resistance in muscle. Journal of Endocrinology, 220(2): 61-79.
Urschel, K. L., & Lawrence, L. M. (2013). Chapter - 6 Amino acids and protein. In: Geor, R.J., Harris, P.A., & Coenen, M., (Eds.). Equine Applied and Clinical Nutrition. Saunders Elsevier: China.
Waller, A. P., & Lindinger, M. I. (2010). Nutritional aspects of post exercise skeletal muscle glycogen synthesis in horses: A comparative review. Equine Veterinary Journal, 42(3): 274-281.
Warren, L. K., & Vineyard, K. R. (2013). Chapter - 7 Fat and fatty acids. In: Geor, R.J., Harris, P.A., & Coenen, M., (Eds.). Equine Applied and Clinical Nutrition. Saunders Elsevier: China.
Wilcox, G. (2005). Insulin and Insulin Resistance Gisela. Alimentary Pharmacology and Therapeutics, Supplement, 22(2): 63-82.
